 A hypothetical landscape aligned vertically, with patterns of conditions changing over time. The variation across the landscape in space and time can be partitioned into pure spatial components, pure temporal components, and a spatio-temporal component. The spatio-temporal variation often has surprising consequences for population growth.
A hypothetical landscape aligned vertically, with patterns of conditions changing over time. The variation across the landscape in space and time can be partitioned into pure spatial components, pure temporal components, and a spatio-temporal component. The spatio-temporal variation often has surprising consequences for population growth.
Causes and Consequences of Spatio-Temporal Heterogeneity For Human Pathogens
The relevant environments for human pathogens are typically the human host, alternative hosts, and the broader ecosystem. Humans are highly heterogeneous in immune status, age, spatial distribution, and behavior. These differences between humans can cause an enormous amount of heterogeneity in the environment relevant to pathogen fitness and spread. For example, we showed that the timing of non-pharmaceutical interventions (NPIs) across different geographic regions could facilitate the fitness and spread of covid-19.
Spatio-temporal heterogeneity can have important, and often unappreciated, effects of the growth rate in spatially distributed populations. We are working to uncover these effects for human pathogens specifically, and for species more generally.
One question is how human behavior responds to variation in the risk of infection. In collaboration with Robert Holt at the University of Florida and Nick Ruktanonchai, and Omar Saucedo at Virginia Tech, we are beginning to work on a new project to model human behavior in response to infection risk. A key question is how human behavior enhances or buffers heterogeneity in the risk of infection for a focal pathogen and for other circulating pathogens. Answering this question relies on being able to accurately model the feedbacks between infection and behavior.
Relevant Publications
Kortessis, N., M.W. Simon, M. Barfield, G.E. Glass, B.H. Singer, and R.D. Holt. 2020. The interplay of movement and spatiotemporal variation in transmission degrades pandemic control. Proceedings of the National Academy of Sciences, 117(48) 30104-30106.
Kortessis, N., G. Glass, A. Gonzalez, N.W. Ruktanonchai, M.W. Simon, B. Singer, and R.D. Holt. In review. Neglected consequences of spatio-temporal heterogeneity and dispersal: Metapopulations, the inflationary effect, and real-world consequences for public health. (preprint)
The relevant environments for human pathogens are typically the human host, alternative hosts, and the broader ecosystem. Humans are highly heterogeneous in immune status, age, spatial distribution, and behavior. These differences between humans can cause an enormous amount of heterogeneity in the environment relevant to pathogen fitness and spread. For example, we showed that the timing of non-pharmaceutical interventions (NPIs) across different geographic regions could facilitate the fitness and spread of covid-19.
Spatio-temporal heterogeneity can have important, and often unappreciated, effects of the growth rate in spatially distributed populations. We are working to uncover these effects for human pathogens specifically, and for species more generally.
One question is how human behavior responds to variation in the risk of infection. In collaboration with Robert Holt at the University of Florida and Nick Ruktanonchai, and Omar Saucedo at Virginia Tech, we are beginning to work on a new project to model human behavior in response to infection risk. A key question is how human behavior enhances or buffers heterogeneity in the risk of infection for a focal pathogen and for other circulating pathogens. Answering this question relies on being able to accurately model the feedbacks between infection and behavior.
Relevant Publications
Kortessis, N., M.W. Simon, M. Barfield, G.E. Glass, B.H. Singer, and R.D. Holt. 2020. The interplay of movement and spatiotemporal variation in transmission degrades pandemic control. Proceedings of the National Academy of Sciences, 117(48) 30104-30106.
Kortessis, N., G. Glass, A. Gonzalez, N.W. Ruktanonchai, M.W. Simon, B. Singer, and R.D. Holt. In review. Neglected consequences of spatio-temporal heterogeneity and dispersal: Metapopulations, the inflationary effect, and real-world consequences for public health. (preprint)
Fragmented landscapes convert suitable habitat (of potentially variable quality) to something less than suitable. Scale transition theory can parse the effects of the matrix locally according to a "mean field" model and the effects of spatial variation.
Matrix effects in fragmented landscapes
Natural populations are not homogeneous across space. Patchiness and heterogeneity are the norm. Humans are large contributors to this heterogeneity and patchiness in space. Clear-cutting forests, urban development, and agricultural land use converts a natural landscape into one where the amount of natural habitat is reduced and highly fragmented. The lost habitat is often unsuitable for many species. What is left behind is often called "the matrix".
While the matrix is generally considered to be unsuitable, that does not mean it is completely inhospitable to life. The matrix can be of varying quality, and have varying effects on population dynamics within patches and dispersal of individuals between patches. What are the effects of the matrix beyond the loss of habitat? Can we mitigate the negative effects of habitat loss through management of the matrix? For what kinds of species and landscapes will managing for the matrix be successful?
In collaboration with Robert Holt, Robert Fletcher, and Emilo Bruna, I am developing models to answer some of these questions.
One thorny issue is the issue of scale. The matrix effects local dynamics within patches and the connections of patches through the dispersal process. We have been using techniques from scale transition theory to understand the joint effects of local and regional effects of the matrix at the entire population scale.
Scaling issues can be quite complicated and yield surprising results. See, for example, some work highlighting surprising scaling effects relevant to disease eradication.
Relevant Publications
Fletcher Jr., R. J., T.A.H. Smith, N. Kortessis, E.M. Bruna, and R.D. Holt. 2023. Landscape experiments unlock relationships among habitat loss, fragmentation, and patch-size effects. Ecology 104(5): e4037
Fletcher, R, J. Sefair, N. Kortessis, R. Jaffé, R.D. Holt, E. Robertson, S. Duncan, A. Marx, and J. Austin. 2022. Extending isolation by resistance for interpreting genetic connectivity across landscapes. Methods in Ecology and Evolution. 13: 2463-2477.
Kortessis, N., M.W. Simon, M. Barfield, G.E. Glass, B.H. Singer, and R.D. Holt. 2020. The interplay of movement and spatiotemporal variation in transmission degrades pandemic control. Proceedings of the National Academy of Sciences, 117(48) 30104-30106.
Natural populations are not homogeneous across space. Patchiness and heterogeneity are the norm. Humans are large contributors to this heterogeneity and patchiness in space. Clear-cutting forests, urban development, and agricultural land use converts a natural landscape into one where the amount of natural habitat is reduced and highly fragmented. The lost habitat is often unsuitable for many species. What is left behind is often called "the matrix".
While the matrix is generally considered to be unsuitable, that does not mean it is completely inhospitable to life. The matrix can be of varying quality, and have varying effects on population dynamics within patches and dispersal of individuals between patches. What are the effects of the matrix beyond the loss of habitat? Can we mitigate the negative effects of habitat loss through management of the matrix? For what kinds of species and landscapes will managing for the matrix be successful?
In collaboration with Robert Holt, Robert Fletcher, and Emilo Bruna, I am developing models to answer some of these questions.
One thorny issue is the issue of scale. The matrix effects local dynamics within patches and the connections of patches through the dispersal process. We have been using techniques from scale transition theory to understand the joint effects of local and regional effects of the matrix at the entire population scale.
Scaling issues can be quite complicated and yield surprising results. See, for example, some work highlighting surprising scaling effects relevant to disease eradication.
Relevant Publications
Fletcher Jr., R. J., T.A.H. Smith, N. Kortessis, E.M. Bruna, and R.D. Holt. 2023. Landscape experiments unlock relationships among habitat loss, fragmentation, and patch-size effects. Ecology 104(5): e4037
Fletcher, R, J. Sefair, N. Kortessis, R. Jaffé, R.D. Holt, E. Robertson, S. Duncan, A. Marx, and J. Austin. 2022. Extending isolation by resistance for interpreting genetic connectivity across landscapes. Methods in Ecology and Evolution. 13: 2463-2477.
Kortessis, N., M.W. Simon, M. Barfield, G.E. Glass, B.H. Singer, and R.D. Holt. 2020. The interplay of movement and spatiotemporal variation in transmission degrades pandemic control. Proceedings of the National Academy of Sciences, 117(48) 30104-30106.
 Winter annual plants in the Chihuahuan Desert of southeastern Arizona. The yearly patterns of seed germination in these species are highly species-specific and contribute to temporal niche partitioning via the storage effect. I ask how such germination responses evolve in in response to temporal environmental variation and competition.
Winter annual plants in the Chihuahuan Desert of southeastern Arizona. The yearly patterns of seed germination in these species are highly species-specific and contribute to temporal niche partitioning via the storage effect. I ask how such germination responses evolve in in response to temporal environmental variation and competition.
Evolutionary robustness of stable coexistence mechanisms.
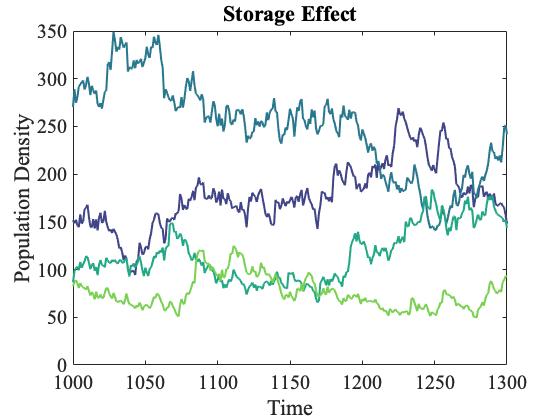
A persistent question in ecology is "why are there so many species?" G. E. Hutchinson famously asked this question in 1959 and those of us in community ecology have been attempting to answer this question ever since. Currently, a large number of stable coexistence mechanisms are hypothesized to allow species to coexist in nature. Coexistence mechanisms delay extinction of species in the presence of competitors, and so are a crucial aspect of standing species diversity. Coexistence mechanisms are all about defining the differences between species that allow species to persist in the presence of each other. Unfortunately, we know little of the origin of the differences required of most coexistence mechanisms and whether such differences persist in the face of evolutionary change. That is to say, are coexistence mechanisms robust to natural selection?
I use the storage effect as a representative coexistence mechanism to study this question for two reasons. First, the requirements for the storage effect to promote coexistence are analogous to the requirements of many other known coexistence mechanisms such as resource and natural enemy partitioning. Second, the theory of the storage effect provides quantifiable descriptions of these requirements. With well developed ecological theory, we can begin to probe how the ecology of coexistence mechanisms influences the evolution of species. Previous focus has been on natural selection, where we can track these quantities as natural selection proceeds. Understanding the evolutionary robustness of species coexistence will hopefully advance our understanding of how communities are constructed and maintained. But there are many other issues to tackle, such as the effect of the genetic and phenotypic composition of species on the selective process itself.
The papers below illustrate these ideas. In them, you can find the components of the storage effect, how they influence selection, and how they evolve as populations adapt in fluctuating environments.
Kortessis, N. and P. Chesson. 2019. Germination variation facilitates the evolution of seed dormancy when coupled with seedling competition. Theoretical Population Biology 130:60-73.
Kortessis, N. and P. Chesson. 2021. Character displacement in the presence of multiple trait differences: Evolution of the storage effect in germination and growth. Theoretical Population Biology 140:54-66.
Kortessis, N. 2021. Digest: The complex interplay of phenotypic variation and diversifying selection. Evolution.
A persistent question in ecology is "why are there so many species?" G. E. Hutchinson famously asked this question in 1959 and those of us in community ecology have been attempting to answer this question ever since. Currently, a large number of stable coexistence mechanisms are hypothesized to allow species to coexist in nature. Coexistence mechanisms delay extinction of species in the presence of competitors, and so are a crucial aspect of standing species diversity. Coexistence mechanisms are all about defining the differences between species that allow species to persist in the presence of each other. Unfortunately, we know little of the origin of the differences required of most coexistence mechanisms and whether such differences persist in the face of evolutionary change. That is to say, are coexistence mechanisms robust to natural selection?
I use the storage effect as a representative coexistence mechanism to study this question for two reasons. First, the requirements for the storage effect to promote coexistence are analogous to the requirements of many other known coexistence mechanisms such as resource and natural enemy partitioning. Second, the theory of the storage effect provides quantifiable descriptions of these requirements. With well developed ecological theory, we can begin to probe how the ecology of coexistence mechanisms influences the evolution of species. Previous focus has been on natural selection, where we can track these quantities as natural selection proceeds. Understanding the evolutionary robustness of species coexistence will hopefully advance our understanding of how communities are constructed and maintained. But there are many other issues to tackle, such as the effect of the genetic and phenotypic composition of species on the selective process itself.
The papers below illustrate these ideas. In them, you can find the components of the storage effect, how they influence selection, and how they evolve as populations adapt in fluctuating environments.
Kortessis, N. and P. Chesson. 2019. Germination variation facilitates the evolution of seed dormancy when coupled with seedling competition. Theoretical Population Biology 130:60-73.
Kortessis, N. and P. Chesson. 2021. Character displacement in the presence of multiple trait differences: Evolution of the storage effect in germination and growth. Theoretical Population Biology 140:54-66.
Kortessis, N. 2021. Digest: The complex interplay of phenotypic variation and diversifying selection. Evolution.
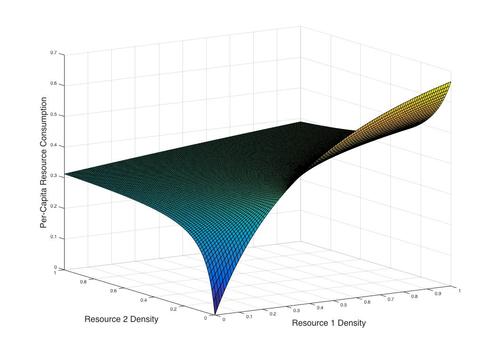 Holling's Type II Functional Response for two resources. Interestingly, increasing the density of one resource can reduce consumption of another.
Holling's Type II Functional Response for two resources. Interestingly, increasing the density of one resource can reduce consumption of another.
Exploitative competition and responses of competitors to resource density.
Competition for consumable resources is prevalent in nature. When species are more dependent on different sets of resources for growth and reproduction, those species may stably coexist via resource partitioning. However, the general requirements for resource partitioning are only well understood when species take a constant fraction of available resources independent of resource density. Both predators (consumers more generally) and their prey have many behaviors that would make these relationships vary with density. Predator searching behaviors, prey anti-predator behaviors, and properties of abiotic resource may vary the amount of resources a single consumer can consume in a given amount of time. The way such nonlinear responses to resource density affect resource competition are poorly understood. Further, current conceptual tools (like R*s) are insufficient to deal with the nonlinear responses to resource density.
Kortessis, N. and CT Lee. In prep. The way consumers respond to resource density determines stabilizing and equalizing effects on coexistence.
Competition for consumable resources is prevalent in nature. When species are more dependent on different sets of resources for growth and reproduction, those species may stably coexist via resource partitioning. However, the general requirements for resource partitioning are only well understood when species take a constant fraction of available resources independent of resource density. Both predators (consumers more generally) and their prey have many behaviors that would make these relationships vary with density. Predator searching behaviors, prey anti-predator behaviors, and properties of abiotic resource may vary the amount of resources a single consumer can consume in a given amount of time. The way such nonlinear responses to resource density affect resource competition are poorly understood. Further, current conceptual tools (like R*s) are insufficient to deal with the nonlinear responses to resource density.
Kortessis, N. and CT Lee. In prep. The way consumers respond to resource density determines stabilizing and equalizing effects on coexistence.
|
Saguaro Pollination Biology
The saguro (Canegia gigantea) is the characteristic plant of the Sonoran desert. It produces large showy flowers during the fore summer drought. The flowers seem to be characteristic of bat pollination. They are white, open at night, located high above the ground, produce copious amounts of nectar, and are fragrant. While other columnar cacti in the Sonoran desert have similar floral phenology and flower characteristics, none have distributions as far north as the saguaro. In the very northern ranges of the saguaro, bats are transient visitors in the summer and may be unreliable pollinators. This hypothesis is corroborated by the flowering phenology of the saguaro. While the flowers open at night, they remain open and are visited by a suite of birds and insects the following morning. In collaboration with Larry Venable and other graduate students in the Ecology and Evolutionary Biology department at the University of Arizona, we are conducting pollinator exclusion experiments to quantify the effectiveness of large and small nocturnal and diurnal pollinators of the saguaro in the northern range of their distribution. |

Saguaro flowers have characters that seem to make them obviously pollinated by bats. The flowers are white, open at night, have copious amounts of nectar, are quite fragrant and are positioned high above the ground. Even though similar species across the range of saguaro distribution are pollinated by bats, there are other pollinators that visit the flowers when they stay open during the day.
|
 Gulf fritillary caterpillar (Agraulis vanillae) consuming a passionflower (Passiflora sp.). Generalist and specialist herbivores are common at our site and can cause substantial damage to plants, even defoliating some individuals.
Gulf fritillary caterpillar (Agraulis vanillae) consuming a passionflower (Passiflora sp.). Generalist and specialist herbivores are common at our site and can cause substantial damage to plants, even defoliating some individuals.
Effects of specialist herbivores on old field community composition
In collaboration with members of the Underwood and Inouye labs at Florida State University, we studied whether a specialist herbivores of an early successional plant affects community diversity and composition. We tracked changes in the cover and composition of an old field community after 5 years of continuous removal of herbivores on Solanum carolinense. Interestingly, we found that specialist herbivores, even on a relatively uncommon plant in the community, affected larger community patterns, suggesting that consideration for less common species -- in addition to dominant species -- are needed for understanding the determinants of community structure.
Kim, T.N., B.J. Spiesman, A.L. Buchanan, A.S. Hakes, S.L. Halpern, B.D. Inouye, A.L. Kilanowski, N. Kortessis, D.W. McNutt, A.C. Merwin, and N. Underwood. 2015. Selective manipulation of a non-dominant plant and its herbivores affects an old-field plant community. Plant Ecology. 216:1029-1045.
In collaboration with members of the Underwood and Inouye labs at Florida State University, we studied whether a specialist herbivores of an early successional plant affects community diversity and composition. We tracked changes in the cover and composition of an old field community after 5 years of continuous removal of herbivores on Solanum carolinense. Interestingly, we found that specialist herbivores, even on a relatively uncommon plant in the community, affected larger community patterns, suggesting that consideration for less common species -- in addition to dominant species -- are needed for understanding the determinants of community structure.
Kim, T.N., B.J. Spiesman, A.L. Buchanan, A.S. Hakes, S.L. Halpern, B.D. Inouye, A.L. Kilanowski, N. Kortessis, D.W. McNutt, A.C. Merwin, and N. Underwood. 2015. Selective manipulation of a non-dominant plant and its herbivores affects an old-field plant community. Plant Ecology. 216:1029-1045.
